�����A�n�����g���ɂ���ĐX�т̊��͑傫���ω�����ƍl�����܂��B ���̕ω��ɂ���Ă������̎��̎�͐��ނ��X�т��r�p���邩������܂���B ����āA�X�т̎p���ǂ̂悤�ɕω�����̂����炩���ߗ\�����Ă������Ƃ���ł��B ���̂��߂ɂ͐X�т��\�������Ȏ�̐����I�Ȑ����ɂ��Ă悭�m���Ă����K�v������܂��B
�����̓�����m�邽�߂ɂ͑��̎�Ɣ�r���邱�Ƃ��L���ł��B �����ŁA����܂ł̌������ʂ����Ƃɔ�r�����݂��Ă��܂����B �������A������n�A������@�Ȃǂ̏������������ƂɈقȂ��Ă���A ������P���ɔ�r���邾���ł͎�ɂ��Ⴂ�m�ɂ��邱�Ƃ͂ł��܂���ł����B �����ŁA�����ꏊ�ň�Ă�������̎��ɂ��Ĕ�r���邱�ƂɂȂ�܂����B
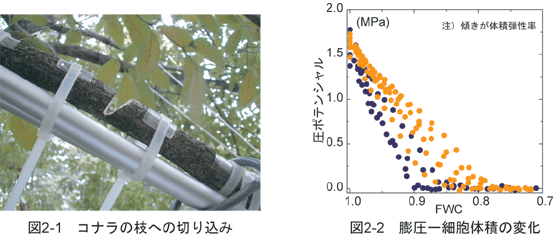
�R�����X�ё����̓c��������ނ̗��t�L�t����c���ň�ĂĂ��܂����B ���̒�����~�Y���A�N���A�~�Y�L�A�R�u�V�A�z�I�m�L�A�~�Y�i���A�R�i���A�P���L�𗘗p���܂����B ����������{�̗≷�ї��t�L�t���тł悭��������ł��B ������U���Ɉ�����U�N���̎����Ƃ�܂��悤�ɑ����g�݁A�����㕔�̗t�𑪒�ł���悤�ɂ��܂����B �^�Ă̐����ԂɈ����ʂ��Č��������x����U���x�Ȃǂ̗t�̐����I�������ɑ��肵�܂���
���̌��ʁA�t�̐����I�����͈���̊Ԃɑ傫���ω����A �Ƃ��ɏ��U���x�̍ő�l�͎�ɂ���đ傫������Ă��܂����i�}1-1�j�B �����ŁA�ǂ̂悤�Ȃ����݂ōő�̏��U���x�ɈႢ�����܂��̂���͂��܂����B ���U���x�iE�j�͓y�납��t�܂ł̐����ʓ��x�iKsoil-leaf�j�ƁA �y��Ɨt�Ƃ̐��|�e���V�����̍��iΨsoil - Ψleaf�j�Ƃɂ���Č��܂�܂��iIshida et al. 1992�j�B
E = Ksoil-leaf �~ (Ψsoil - Ψleaf)
��͂̌��ʁA�����ʓ��x�͎�ԂŗL�ӂȍ�������܂���ł����B ����ŁA���|�e���V�������͎�ɂ���ėL�ӂɈقȂ�A �ő�̏��U���x���傫����قǑ傫���X��������܂����B ����ɁA�R���������̒��삳��̎����������肵�A�t��PV�Ȑ��i�E���Ȑ��j�𑪒肵�܂����B ���̋Ȑ�����t�̐��|�e���V�������ǂ̂��炢�ቺ�����Ƃ��ɁA�זE�̖c����������̂��i������_�GΨtlp�j�]�����邱�Ƃ��ł��܂��B ���̌��ʁA�����̐��|�e���V�����̍����傫����قǁA������_���Ⴂ���Ƃ��킩��܂����i�}1-2�j�B
���������āA�����ɍő�̏��U���x�̑傫����́A�t�Ɠy��̊Ԃ̐��|�e���V�����̍����傫���A ����́A�t�̐��|�e���V�������ቺ���Ă��c�����ێ��ł��邩�炾�Ƃ������Ƃ��킩��܂����B ���̌����̌��ʂ́A�t�̐������������ؑS�̂̐����̗A���ɑ傫���e�����邱�Ƃ������Ă��܂��B �����ŁA�t�̐��������ɂ��Ă���ɒ��ׂ邱�Ƃɂ��܂����B
�t�̐����Ɋւ��鐫���ɂ��ẮA�Ƃ�킯�t�̍זE�̖c����ۂ����݂��d�v�ł��B �c���������ƁA��ӂȂǂ̍זE�̐����I�����ɏ�Q������邩��ł��B �����������ł͖c���������₷���̂ŁA�t�̋z���\�͂����シ�邱�Ƃɂ���Ėc���ێ��\�͂����߂�܂��B �t�̋z���\�͂́A�זE���ɗn�����~�ς��A�זE���t�̐Z���|�e���V�������ቺ���邱�ƂŌ��サ�܂��B ���̍זE�̓�����Z�����߂Ƃ����A�A�����鐅�X�g���X�Ƃ̊W���ڂ������ׂ��Ă��܂����B
����ŁA�זE�̖c���ێ����l����Ƃ��ɂ́A�c���̗��������厖�ł��B ���Ȃ킿�A�זE�������������ʂɑ��āA�ǂ̂��炢�c�����ቺ���邩�Ƃ������Ƃ��l����K�v������܂��B �������A����܂ł��̐����i�̐ϒe�����j�ɂ��Ă͏\���Ɍ�������Ă��܂���ł����B �����ŁA����w�̎����搶�̉��ő̐ϒe�����ɂ��ďڂ������ׂ邱�Ƃɂ��܂����B
���@�ڎ���