| 植物生態学研究室のメンバーのページへ |
低リン酸環境に対するルピナスの応答機構の解明舟山(野口)幸子 [PD] |
| これまでの私の研究を紹介します。大学院時代からポスドク初期は、ウイルスに感染したヒヨドリバナの研究をしていました。このときは、葉やシュートといった植物の地上にある部分を主に調べていました。その後はいろいろなご縁があり、根とリン栄養の研究をはじめて今に至っています。"The hidden half”とも言われる植物の根はわかっていないことも多く、根の研究はとても面白いです。 |
| (1) 植物の根とリン栄養に関する研究 |
| (2) ジェミニウイルスに感染したキク科ヒヨドリバナ Eupatorium makinoi に関する研究 |
| (3) その他の研究(主に共同研究) |
1.植物の根とリン栄養に関する研究 | ||||||||||
| リンは、植物にとって重要な必須元素です。しかし多くの土壌では、植物が吸収できるリン(水に溶けているリン酸)の濃度は低いことがわかっています。そのため、野外の植物は、少ないリンを効率よく吸収・利用するためにさまざまな性質を持っています。 | ||||||||||
| 農業で使われるリン肥料は、リン鉱石から作られます。地球上のリン鉱石は有限で、遠くない未来に枯渇すると推測されています。低リン環境に生育する野外植物のリンの吸収・利用方法を明らかにすることは、少ないリン肥料でもよく育つ作物の育種に役に立つ可能性もあります。 | ||||||||||
植物の根は土の中にあり目にすることは少ないですが、驚くほどに多様な形をしていて、さまざまな機能を持っています。私は、土の中で不足しがちなリンを植物の根がどのように上手く獲得しているのかを研究しています。

1.1 ヤマモガシ科常緑木本Hakea prostrataのクラスター根の呼吸特性
リンが極端に少ない土壌で有名な西オーストラリアでよく見られるヤマモガシ科植物は、「クラスター根 (cluster roots)」と呼ばれる試験管ブラシのような形の房状の根を作ります。クラスター根は、大量の有機酸(クエン酸、リンゴ酸など)や酸性ホスファターゼを合成・放出し、土の粒子と結合しているリン酸を可溶化して効率よく吸収します。 ヤマモガシ科のHakea prostrataのクラスター根では、ミトコンドリア呼吸電子伝達鎖でATP合成と共役しない末端酸化酵素AOX が誘導されます。これは、有機酸を合成するときに同時にできる大量の還元力 (NADH) を消費して、呼吸電子伝達鎖が過還元状態になるのを防ぐために役立っていると考えられています。 原著論文
1.2 リンによって発現が変化するCLE遺伝子
CLE遺伝子は、植物の形態形成や環境応答で働くペプチドホルモンをコードしています。ミヤコグサ Lotus japonicus は日本全土に自生する草本で、マメ科のモデル植物として微生物共生の研究を中心に多くの研究で使われています。ミヤコグサでは、硝酸(窒素)を与えると根で発現が増えるCLE遺伝子が見つかっています。 そこでリンに着目して調べたところ、ミヤコグサでは、環境のリン濃度が上昇したときに根で発現が著しく増加するCLE遺伝子が2つ見つかりました。これらのCLE遺伝子は、植物の根が環境のリン濃度を感知する機構に関わっている可能性があるかもしれません。 原著論文
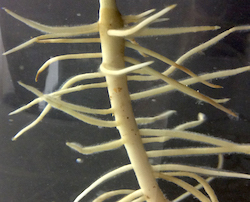
1.3 リン欠乏に対するルピナス2種の応答機構(現在継続中)
マメ科のシロバナルピナス Lupinus albus はリン欠乏に強く、クラスター根を作ります。近縁のホソバルピナス L. angustifolius もリン欠乏に強いのですが、クラスター根は作りません。現在私は、根の形が異なるシロバナルピナスとホソバルピナスのリン欠乏応答を調べています。 クラスター根は、効率的なリン吸収に役立つと考えられています。植物では器官(根、茎、葉など)を作るのにエネルギーが必要で、そのエネルギーのことを構成コストと呼んでいます。シロバナルピナスはリン欠乏下で構成コストが高いクラスター根を作るのに対し、ホソバルピナスは構成コストが小さい根をリン欠乏下で大量が作っていました。これは、ホソバルピナスが根をたくさん作ることでリンの吸収量を増やし、リン欠乏を乗り切っていることを示唆しています。 シロバナルピナスとホソバルピナスは、リン欠乏下の成長速度がほぼ同じです。植物の成長は、光合成で獲得した炭素(収入)と呼吸や有機酸放出などで失った炭素(支出)のバランス(個体の炭素収支)と密接に関係しています。リンが欠乏下で、シロバナルピナスは呼吸速度と有機酸放出速度が高いクラスター根を作ったのに対し、ホソバルピナスは呼吸速度と有機酸放出速度が低い根を大量に作りました。光合成、呼吸、有機酸放出の測定値を用いて計算すると、2種は同じような個体の炭素収支を示しました。これが2種の成長速度がほぼ同じ理由だと考えています。 原著論文
このページのトップへ |



2.ジェミニウイルスに感染したキク科ヒヨドリバナ Eupatorium makinoi に関する研究 | |||||
| 私は、大学院生のときにジェミニウイルスに感染したヒヨドリバナの研究を始めました。病気は、多くの種が一緒に生えている野外の状況において、特定の種(宿主植物)の特定の遺伝子型(罹病性植物)の個体にだけ悪い影響を及ぼし、感染個体と周囲の植物との力関係を変化させます。このことから、植物の病気は生態系において重要な役割を持っていると考えられています。 | |||||
研究を進めるうちに、ウイルス感染して葉脈の周りが黄色くなったヒヨドリバナの葉は、光が十分にあるときの光合成速度があまり低下しないことがわかりました。これは、ウイルスに感染されても受ける影響が小さかった個体が生き残ってきたからかもしれません。今は中断していますが、いつかウイルスに感染した葉が黄色くなる仕組みを明らかにしたいと思っています。

総説
2.1 ウイルス感染したヒヨドリバナの個体群動態
野外では、特に明るい場所で、ウイルスに感染して葉が黄色くなっているのに成長し花を咲かせているヒヨドリバナをよく見かけます。そこで、「ウイルス感染の影響は光環境によって異なる」という仮説をたてて、自生地で8年間の調査を行いました。感染流行後の個体群の衰退速度は光環境によって異なり、明るい場所の個体群はゆっくり衰退することがわかりました。しかしどの個体群もウイルス病流行によって絶滅する可能性がきわめて高いことも明らかになりました。 原著論文
2.2 ウイルス感染がヒヨドリバナの成長に及ぼす影響明るい生育地でヒヨドリバナへのウイルス感染の影響がゆっくり表れた原因を明らかにするために、3段階の光条件下でヒヨドリバナを栽培し、成長解析を行いました。この成長解析では、感染による弱光下の光合成速度の低下が物質生産の減少を介して個体の成長を低下させることを定量的に示しました。 野外では、成長が悪くなって草丈が小さくなると、周囲の植物から被陰されて受光量が少なくなります。その結果、ますます成長が悪くなり、さらに受光量が少なくなります。この負のループが暗い生育地では速く進むために、ヒヨドリバナに対するウイルス感染の影響が強く出たと推測されます。逆に明るい生育地ではもともとの受光量が多いため、ウイルス感染の影響がゆっくり表れたのだと考えています。 原著論文
2.3 ウイルス感染したヒヨドリバナの葉の光合成
ウイルスに感染したヒヨドリバナの葉では、葉脈の周辺が黄色くなります。感染葉では弱光下の光合成速度が顕著に低下したのに対し、飽和光下の光合成速度はほとんど低下していませんでした。感染葉では光吸収に重要な集光アンテナタンパク質が著しく減少し、これが感染葉の光合成低下の主な原因であることが明らかになりました。 感染で葉が黄色くなる仕組みを明らかにするために、「クロロフィル合成が抑制されて集光アンテナタンパク質が減少する」という仮説をたてて解析を進めています(現在は中断中)。 原著論文
このページのトップへ |


3.その他の研究(主に共同研究) |
| 今までいくつか共同研究をさせていただきました。内容は研究業績のページをご覧ください。 |
| おわりに |
| 東京にきてからは、ほとんどずっと植物のリン欠乏応答の研究をしていました。そんな中、思いがけず東京薬科大学で葉脈が黄色くなったヒヨドリバナに再会しました。ヒヨドリバナの研究もまたやりたいなと思っています。 |
| このページのトップへ |
| 植物生態学研究室のページへ |