

Since the pioneering review of Burdon (1987), the interest in the role that pathogens play in non-agricultural plant communities is rapidly expanding. While plant pathogens can often cause highly selective damage to their host plants, they leave adjacent non-host plants untouched. By reducing competitive capacity or killing individual plants, pathogens may affect the population dynamics of host plants, the genetic structure of host populations, the evolution of sexual reproduction, the distribution of plant species, and the diversity of plant communities.
I have been conducting ecological and physiological studies concerning effects of geminivirus infection on Eupatorium makinoi (Asteraceae), and now, study on the role of alternative oxidase (AOX) in the resistance to TMV in tobacco plants. Here, I summarize my studies briefly.
|
|
|
| Fig. 1-1 | Fig. 1-2 | |
| Fig. 1 Study site | ||
Eupatorium makinoi Kawahara et Yahara (Asteraceae) plants are perennial, common in open disturbed places along roadsides or forest margins in Japan. They are frequently infected by tobacco leaf curl geminivirus in their natural habitats. Infected plants are distinguished by a visible symptom, yellow variegation.
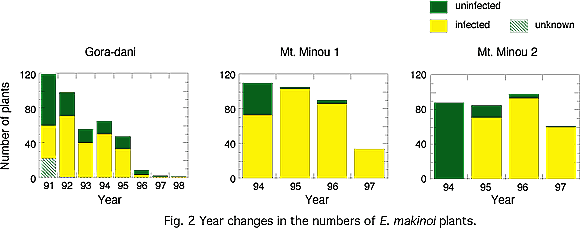
We examined the population dynamics of E. makinoi during virus epidemics in the two field sites, Gora-dani (Fig. 1-1) and Mt. Minou (Fig. 1-2). The Gora-dani population rapidly declined after the virus epidemic to a state of near extinction (Fig. 2). In the Minou populations, individual numbers also decreased during the virus epidemics (Fig. 2), although they did more gradually.
We followed the fate of individual plants in Gora-dani, and showed (1) the adverse effects of infection on growth made E. makinoi plants small, and (2) mortality of infected plants increased with decreasing plant height. Therefore, it can be concluded that the virus-infected plants died due to the adverse effects on growth and size-dependent mortality, and consequently, the population in Gora-dani rapidly declined after the virus epidemic in the field.
The geminivirus infecting E. makinoi is famous for the oldest record of a plant viral disease. In other words, the geminivirus and E. makinoi have coexisted for a long time. However, the local population became extinct after the virus epidemic. These suggest that some mechanisms underlie the long-term coexistence between them. One of the future directions of this project is to clarify the mechanism of the long-term coexistence.
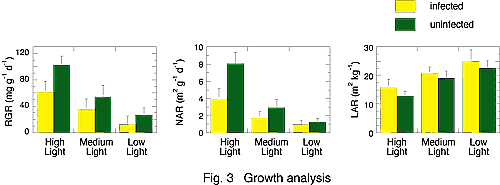
The decrease in growth by virus infection determined the population dynamics of E. makinoi through size-dependent mortality. Then, we conducted growth analysis using virus-infected and uninfected E. makinoi to clarify what component caused the decrease in the growth rate in infected plants. Relative growth rate (RGR) and net assimilation rate (NAR) were significantly decreased by virus infection, whereas leaf area ratio (LAR) was significantly greater than that of uninfected plants. These results suggested a close relationship between RGR and NAR.
We estimated NAR from light-response curves of leaf photosynthesis, in situ measurements of irradiance, and respiration rates of leaves, stems, and below-ground parts. The estimated NAR agreed well with the NAR obtained by conventional growth analysis techniques. The decrease in this estimated NAR in infected plants were mostly explained by a decrease in leaf photosynthesis. Thus, we concluded that the lowered RGR in virus-infected plants was attributed mainly to the impaired photosynthesis in leaves.
 |
|
| Infected | Uninfected |
| Fig.4 Eupatorium makinoi |
|
 |
|
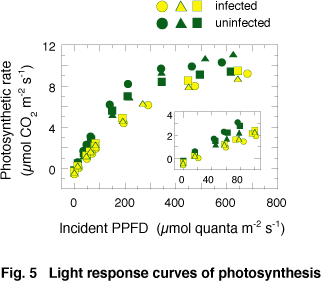
It is important to study photosynthetic properties of E. makinoi leaves infected by the geminivirus, because the impaired photosynthesis underlaid the ecological phenomena. Fig. 5 shows typical light-response curves of fully developed leaves. The initial slope of light-response curves of photosynthesis (photosynthetic rate under ambient CO2 condition on a leaf area basis) measured on an incident quantum basis was significantly lower in infected leaves than that in uninfected leaves. On the other hand, the light-saturated photosynthetic rate (Pmax) in infected leaves was comparable to that in uninfected leaves. The decrease in the initial slope was caused by the decrease in light absorptance, which was attributed to the small amount of Chl proteins, particularly LHCII.
In severely yellowed leaves, not only the initial slope, but also Pmax decreased. Since the depression of Pmax occurred only in severely infected leaves, the mechanism of the decreased Pmax would be different from that of the decrease in the initial slope.
Leaf chlorosis was caused by less Chl accumulation in infected leaves. We examined 5-aminolevulinate (ALA) synthesis, which is one of the rate-limiting steps of Chl biosynthesis. ALA-synthesizing capacity was lowered in infected leaves, which suggested that the activity of Chl biosynthesis impaired in infected leaves. Further studies of both plants and viruses at the molecular level may clarify the relationship between virus replication and the impairment of Chl synthesis.
I studied on cluster roots of H. prostrata for 4 months in Perth.