研究テーマ1: 葉の光合成光順化能力の種間差
植物が受ける光環境は時間的、空間的に多様である。これに対し動くことのできない植物は、それぞれの環境において効率よく光合成を行うために様々な性質を変化させていることが知られており、葉レベルから個体・群落レベルまで様々な研究がなされてきた(Pearcy & Sims 1994 in Exploitation of environmental
heterogeneity by plants pp. 145-174)。光順化の一例としては、明るい環境において、厚く、窒素含量が多く、光合成能力の高い陽葉を、暗い環境において、薄く、窒素含量が低く、光合成能力の低い陰葉をつけることが広く知られている(Björkman 1981 in Physiological plant
ecology pp. 57-107)。明るい環境では十分な光があるため、より多くの葉緑体を持つことで高い光合成を実現できるが、暗い環境では十分な光がないため、多くの葉緑体を持ったとしても、下層の葉緑体は十分な光を吸収することができず、コスト当たりの光合成の低下や、呼吸による炭素獲得の低下を引き起こすためと考えられる。しかし、種によっては明るい環境においても光合成能力を高めないものや、かえって低下してしまうものも存在する。この種間差はどのような生理的要因によって生じているのか、そしてその背景にはどのような生態学的意義があるのだろうか。
葉の光合成光順化と解剖学的性質の関係
これまで、陽葉が陰葉よりも厚いのは高い光合成能力を持つために葉面積あたりの葉緑体を多く持つ必要があるためと考えられてきた(Terashima et al. 2001 Journal of Plant
Research 114: 93-105)。葉緑体は、CO2の拡散抵抗を減らすために、細胞表面付近に位置する必要がある。もし細胞表面から離れれば、CO2の供給不足がおき、光合成の効率は著しく低下する。この状況で葉が光合成能力を高めるには、葉緑体を増やすために葉を厚くし細胞表面積を増やす必要がある。しかし、成熟葉の光順化はこの知見と一見矛盾する。弱光で展開が終了した葉を強光へ移すと葉の厚さの増加を伴わずに光合成能力が上昇することが、多くの先行研究で観察されている(Jurik et al. 1979 Plant Physiology 63:
542-547)。これは、光合成能力の増加には葉の厚さの増加は必要ないことを意味するのだろうか。そして、厚さの変えられない葉はどのように光合成能力を高めているのだろうか。
また、弱光で展開した葉が強光にさらされた後に起こる光合成能力の増加が大きいほど新たな光環境での成長促進が大きいと考えられるが、増加の程度は種によって異なる(Turnbull et al. 1993 Oecologia 94:
218-228; Yamashita et al. 2000 Oecologia 125: 412-419)。光順化能力の大きさはどのように決まっているのか。そして、なぜ種によって成熟葉の光順化能力が異なるのか。
これらの疑問に答えるため、私は葉の解剖学的性質に着目した。近年、葉内のCO2拡散など光合成における葉の解剖学的性質の重要性が指摘されている(Evans et al. 1994 Australian Journal of Plant Physiology 21:
475-495)。多くの種において葉の形態はいったん成熟すると環境が変わっても大きく変化することはない(Sims
& Pearcy 1992 American Journal of Botany 79: 449-455)。そこで、葉の解剖学的性質が光合成の光順化能力を制限するのではないかと考え、以下の研究を行った。
(1)葉の光合成光順化に解剖学的性質の変化は必要か?
厚さの変えられない成熟葉が光合成能力を高めるメカニズムを明らかにするため、弱光で生育したシロザの強光への移植実験を行った。移植により光合成能力は11μmol m-2 s-1から17μmol m-2 s-1へと有意に上昇したが、展開以前から強光条件で生育した葉には及ばなかった。ルビスコあたりの光合成能力、葉緑体中のルビスコ濃度、葉緑体の厚さ、葉緑体が細胞間隙に接する面積、葉面積あたりの葉緑体数に分けてこの順化を評価したところ、葉緑体が細胞間隙に接する面積のみが有意に増加していた。つまり、弱光生育の葉では細胞表面を葉緑体が完全に占有してはいなかったため(78%)、強光へ移したあとにその部分を葉緑体が占有するように大きくなることで光合成能力の上昇が起きたのである(図1)。一方で、順化後の葉緑体による細胞表面の占有率は約95 %に達し、それ以上葉緑体を増やす余地はほとんどなく、光合成能力も強光生育の葉には及ばなかった。これらの結果から、陰葉は光環境の向上に順化して光合成能力を上昇させるために、葉を厚くして葉緑体が存在しない隙間をあけていることが示唆された(1: Oguchi, Hikosaka
& Hirose 2003)。
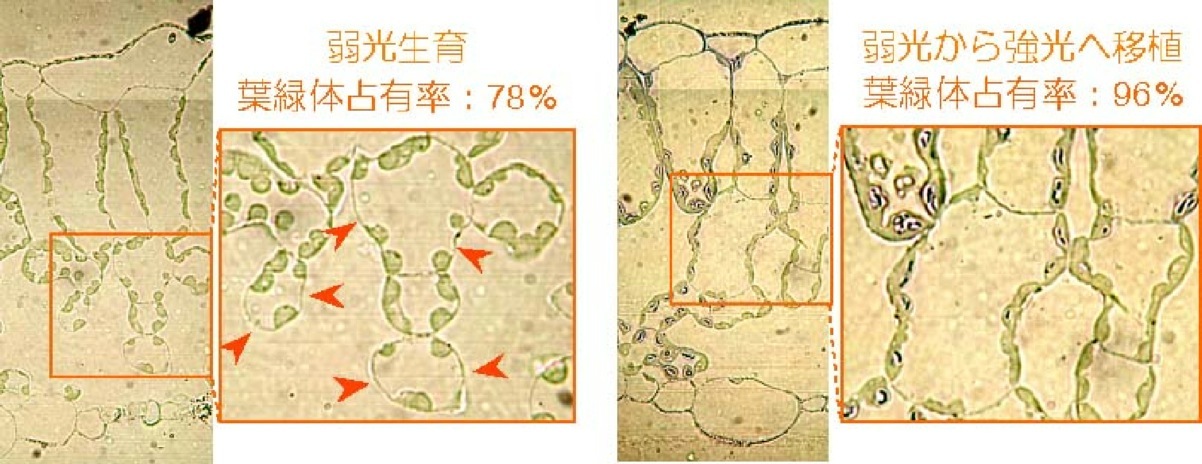
図1 左:弱光生育の葉、右:弱光から強光へ移植した葉の断面写真。矢印は細胞表面で葉緑体の存在しない部分を示す(細胞同士が接している部分は含まない)。弱光生育の葉では海綿状組織を中心に葉緑体が存在しない隙間が見られるが、強光へ移植した葉では細胞表面を葉緑体がほとんど埋めている。
(2)光合成光順化における葉の解剖学的制約:木本三種における光環境の上昇に対する異なった応答
光順化能力が種間で異なるメカニズムを調べるため、遷移系列上での出現時期が異なる木本三種の実生を用いて弱光から強光への移植実験を行った。遷移先駆種であるダケカンバではシロザと同様に葉緑体が大きくなることで光合成能力を増加させていた。遷移中間種であるウリハダカエデは、展開終了後であっても葉を厚くすることで葉緑体を増やすことが可能になり、光合成能力が増加していた。遷移後期種であるブナは、陰葉の細胞表面にほとんど隙間が無く、葉の厚さを変えることもなかったために、光合成能力が増加しなかった。これらの結果から、光順化能力を持つためにはあらかじめ葉を余分に厚くして葉緑体が延びるための隙間を空けておく必要や、光環境が向上した後に葉を厚くするため細胞壁を再構築可能な状態にしておく必要があると考えられた。前者は葉を厚くするためのバイオマスの投資が必要である。また後者は葉の強度を高めたり被食防御を行うことができないと考えられる。このようなコストは、光環境の向上を期待する楽観的な種にとっては払う価値があるかも知れないが、そのような光環境の向上を期待しない悲観的な種にとっては払う必要のないものと考えられる。このような二者択一的な戦略が存在するために、光順化能力に種間差が存在すると考えられた(2: Oguchi, Hikosaka
and Hirose 2005)。
(3)冷温帯樹林におけるギャップ形成に対する木本実生の光合成光順化
森林内部では上層の個体や枝が枯死しギャップが形成されることで突然林床に到達する光が増えるなど、光環境は大きく変動する。実際の自然環境における光合成光順化のメカニズム、種間差の原因を調べるため、冷温帯広葉樹林で林冠木を倒してギャップを形成し、林床に生育する木本実生の葉の応答を観察した(図2)。材料には遷移系列上での出現時期や生活型の異なる4グループ(遷移先駆種、亜高木種、遷移後期種、つる植物)、8種を用いた。ギャップ形成後はつる植物であるツルアジサイ、イワガラミ以外の全ての種で光合成能力が増加した。ハリギリ、ホオノキ、ミズナラではシロザ、ダケカンバと同様に葉緑体が大きくなることで光合成能力が上昇していた。ハウチワカエデ、イタヤカエデではウリハダカエデと同様に葉肉細胞表面積が大きくなることで光合成能力が上昇していた。つる植物は解剖学的な性質の変化は見られなかった。光環境の上昇に対する応答や順化のメカニズムは種間で異なったが、葉緑体が細胞間隙に接する面積の増加が光合成能力の増加に必要であったことは共通していた。このことは自然環境においても葉の解剖学的性質が光合成能力を制限することを意味する(3: Oguchi, Hikosaka,
Hiura and Hirose 2006)。

図2 左:ギャップ形成前の林床の様子 右:ギャップ形成後の林床の様子
(4)ハリギリにおけるギャップ形成に対する光合成光順化のコストとベネフィット
上記の研究により、成熟葉の光順化は光合成能力を高めるが順化能力を持つためにはコストが必要であることが明らかになった。自然環境において、光順化はコスト以上のベネフィットをもたらしているのだろうか。そこで、光合成モデルを用いて自然環境での炭素獲得量を推定した(図3)。材料は、ギャップ形成後に細胞表面の隙間を埋めるように葉緑体を大きくすることで光合成能力が上昇したハリギリを用いた。ギャップ形成後、光合成光順化によって最大炭酸同化速度、最大電子伝達速度、呼吸速度が増加した。ギャップ形成による光環境の向上により炭素獲得量は4.5倍程度にまで増加した。このとき、モデルを用いて感度分析を行ったところ、光合成能力が上昇した葉の炭素獲得量は、光合成能力が上昇しなかった場合よりも11.4%高いことが示唆された。この炭素獲得量の増加は、葉緑体を大きくするための隙間を作るのに投資したバイオマス量よりも大きかった。これらの結果は、ギャップ形成に対する光合成光順化のベネフィットが、順化能力を持つために投資したコストよりも大きかったことを意味する(6: Oguchi, Hikosaka,
Hiura and Hirose 2008)。
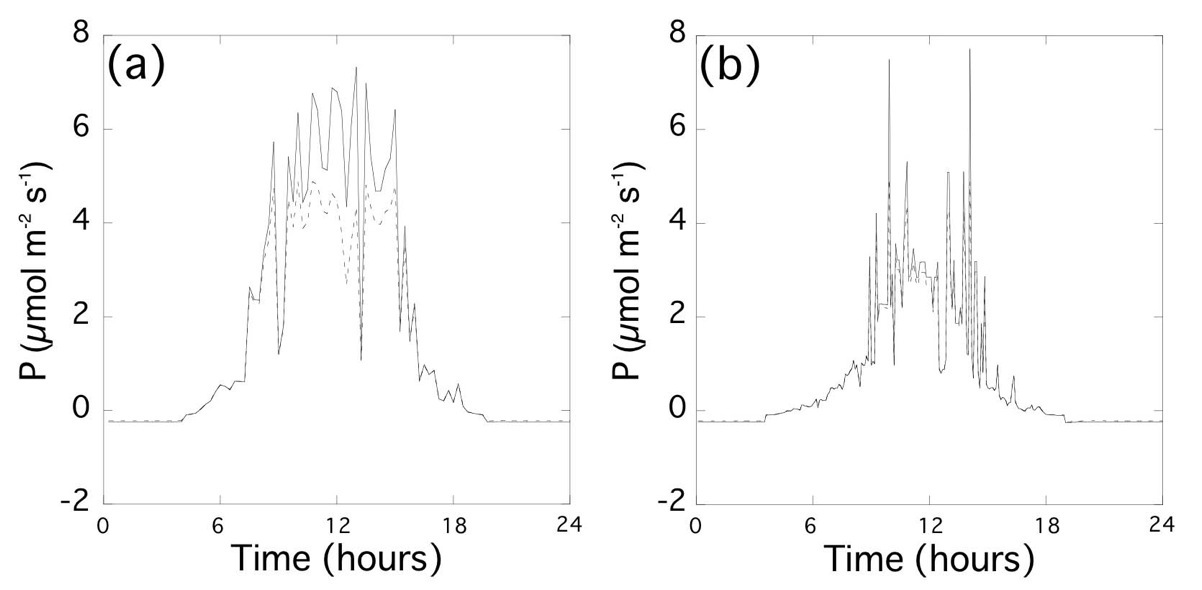
図3 光合成モデルから推定した光合成速度(P)の日変化の一例。ギャップ環境(a)とコントロール環境(b)における、ギャップ生育の実生(実線)とコントロール生育の実生(破線)の光合成速度。実線と破線の差が光順化の影響に当たる。
これまでの研究により、成熟葉の光順化能力は葉の解剖学的性質によって制限され、葉緑体が細胞間隙に接する面積を増やすことが可能な種でのみ、光合成能力の上昇が可能であることが明らかになった。成熟葉の光順化能力を持つためには、葉緑体を伸ばす隙間を空けるために葉を厚くするコストや、葉の強度や病原菌耐性を抑えて厚さの可塑性を保つというリスクを負う必要があると考えられた。また、ギャップ環境での光合成速度を計算したモデルは、光順化による光合成能力の上昇が炭素獲得量を増やすことを示した。しかし、実生がギャップ形成を経験できる確率は不確定であるために、光順化能力を持つためのコストやリスクを抑える戦略を採る種が存在すると考えられ、これが光順化能力の種間差をもたらすものと考えられる。
葉の光合成光順化能力の種間差と成長・生存への影響
(5)冷温帯落葉広葉樹林でのギャップ更新における、木本実生の光合成可塑性と林床生存率とのトレードオフ
(3)で述べたように、ギャップ形成は森林の更新に欠かせないイベントである。光強度の上昇に対する実生の応答は種によって異なっており、森林内での多様性維持に貢献していると考えられている。葉レベルにおいてもギャップ形成に対して光合成能力の応答が起こる。この応答は(1)から(4)で紹介したような既に展開していた葉の順化だけでなく、新しく展開する葉の性質を変えることによっても起こり、どちらも種間差があることが知られている。
光合成能力の可塑性の種間差は、ギャップ形成後の実生の成長に寄与しているのだろうか? 寄与しているとすれば、光合成能力の可塑性の種間差も種の共存に貢献していると考えられる。また、なぜ種によっては光合成の可塑性を持たないのだろうか?
光合成の可塑性にはなんらかのコストが必要であり、林床での生存に負の影響を与えているのだろうか?
これらの疑問に答えるため、ギャップ形成後の6種の木本の光合成と成長の応答を追跡した。ギャップ形成後4年間の実生の相対成長速度は、ギャップ形成後に展開した葉の光合成能力と強い相関を示した(図4左)。また、相対成長速度と光合成能力のギャップ応答の大きさは、8年後の生存率と負の相関を示した(図4右)。このことは、ギャップ応答能力と林床での生存能力との間にトレードオフの関係があることを示唆している。この関係により、種によっては林床での生存を優先するため、光合成の可塑性を持たないことが考えられる。この種間の戦略の違いがギャップ更新を介して森林内での種の共存に貢献していると考えられる(Oguchi et al. 2017
Tree Physiology: DOI 10.1093/treephys/tpx042)。
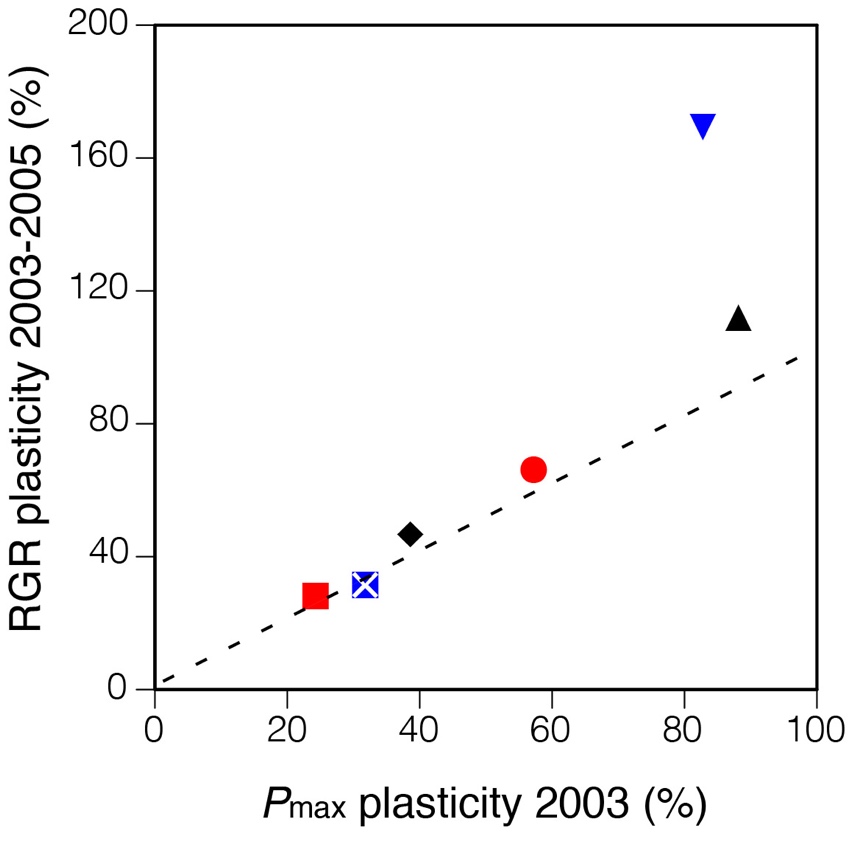

図4 ギャップ形成翌年の光合成能力の可塑性(Pmax plasticity 2003: controlに比べてgapでどれだけ光合成が上昇したか)は、相対成長速度(RGR)のgapでの促進率と正の相関を示した一方(左図)、control(林床)での死亡率と正の相関を示した(右図)。つまり、gapでは早い成長を実現できるが、林床での生存率は低下してしまうというトレードオフが存在することを示唆している。(●ハリギリ、■ホオノキ、◆ハウチワカエデ、▲アオダモ、×イタヤカエデ、▼ミズナラ)