
これまでの主なフィールドサイト 左から、ソルトレークシティー(アメリカ・ユタ州,砂漠気候)、ジャレジャレ・ヒマール(ネパール,高山帯)、縞枯山(長野県茅野市,亜高山帯)
文献: 種子田ら(2016)日本生態学会誌 66:447-464
植物の使う水は、土壌から根で吸収され、植物体内を通って蒸散している葉へと運ばれます。土壌への水は降雨などによってもたらされますが、安定して供給されるとは限りません。さらに、植物体内の根から葉までの移動経路にあたる組織は、必ずしも水をよく通すわけではありません。このため、乾燥地はもちろん日本のように湿潤な気候に生える植物でも、個体内の水の移動効率や利用の仕方が植物の成長やかたちを制限する重要な要因になります。
私たちは植物体内の水の移動の様子や水の使われ方を詳しく調べていくことで、植物のかたちや適応戦略の進化を明らかにしています。こうした研究を通して、陸上環境における植物の適応放散の一端を明らかにするとともに、地球の環境変動に対する植物の応答の予測に役立つ情報を提供したいと思っています。
これまでの主なフィールドサイト 左から、ソルトレークシティー(アメリカ・ユタ州,砂漠気候)、ジャレジャレ・ヒマール(ネパール,高山帯)、縞枯山(長野県茅野市,亜高山帯)
文献: 種子田ら(2016)日本生態学会誌 66:447-464
私たちは、生活史戦略の異なる植物種間のおける道管や仮道管のかたちや水輸送機能の違いがどのように環境適応にかかわるのかに興味を持って研究を行っています。
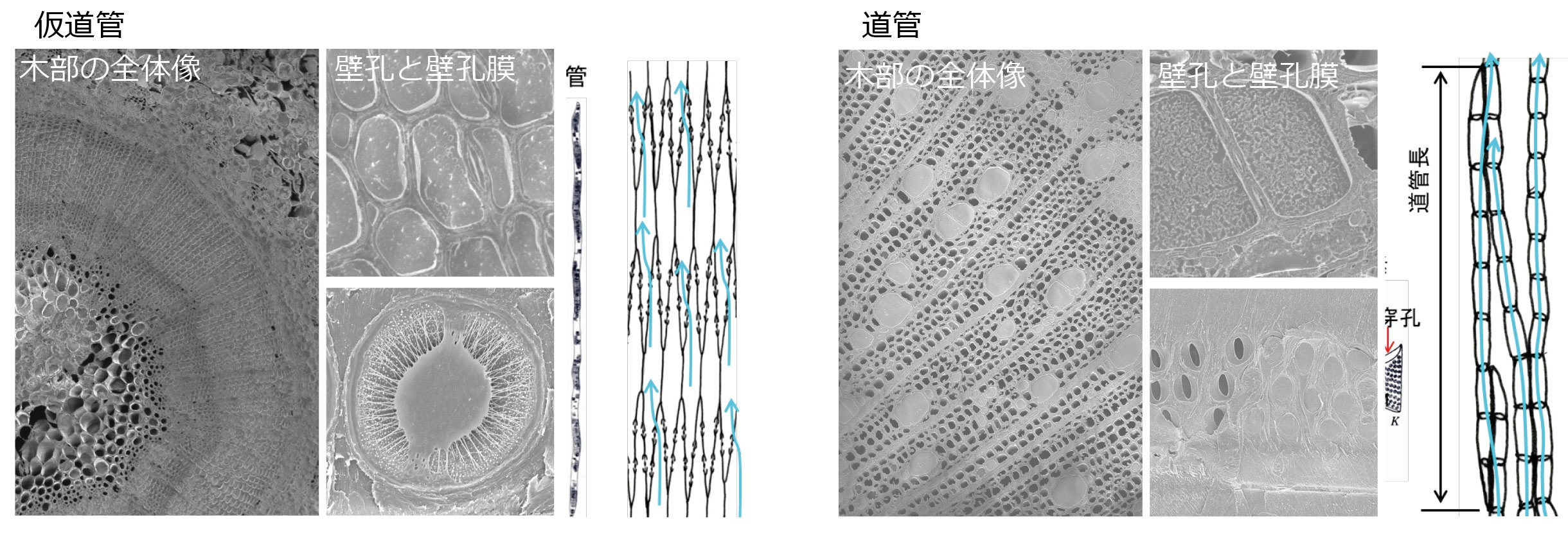
仮道管と道管のかたちと水の流れ方
文献: Ooeda H et al. (2018) Tree Physiol 38:223-231; Yazaki K et al. (2020) Amer J Bot (印刷中)
私たちは、根、茎、葉の間のバイオマスの分配パターンに注目して、植物のデザインを決める機能や環境要因について考えています。これまでの私たちの研究から茎では水輸送機能よりも力学的な支持機能が重要であり、根では水と窒素の両方の資源の吸収が最適なるようにバランスが決まると予測されています。
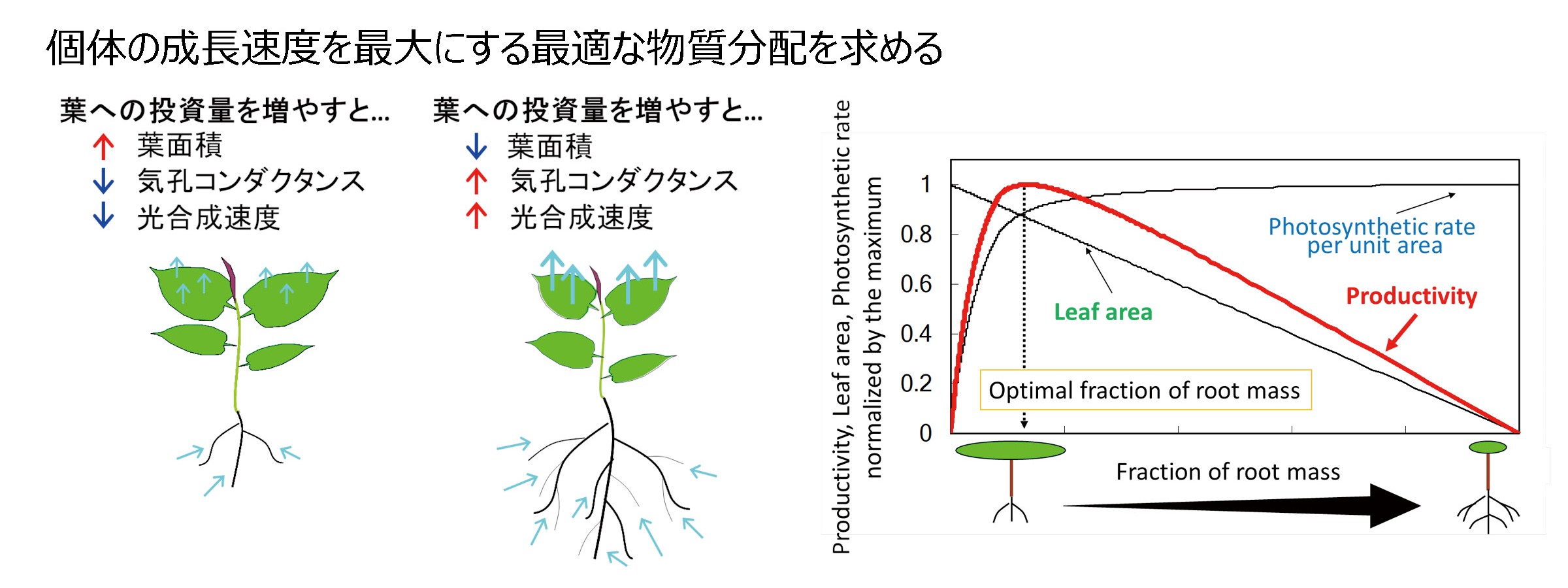
成長を最適にする物質分配
文献: 種子田、舘野(2003)生物科学 54:154-163; Taneda H & Tateno M (2004) Amer J Bot 91:1949-1959
植物体内における水の移動は、電気回路における「オームの法則」と同じルールに従います。そこで、この法則をあてはめて単純に考えると、水の移動距離が長かったり、多くの分枝があったりすると樹冠の先端にある葉ほど水が供給されにくくなることが予想されます。
こうした問題は、ほかの組織にも適用できます。例えば、葉では複雑に分枝した葉脈を通って葉の隅々にまで水が供給されなければなりません。そして、この「輸送距離や分枝による効果」を克服しない限り、葉の周縁部へ水が届かなくなってしまいますし、樹冠では水供給の制約により自由に枝や葉を伸ばすことはできなくなるはずです。
私たちは、植物体内や組織内の水の流れやすさの分布を詳しく調べて、葉身や樹冠の隅々にまで水が供給される仕組みを研究しています。
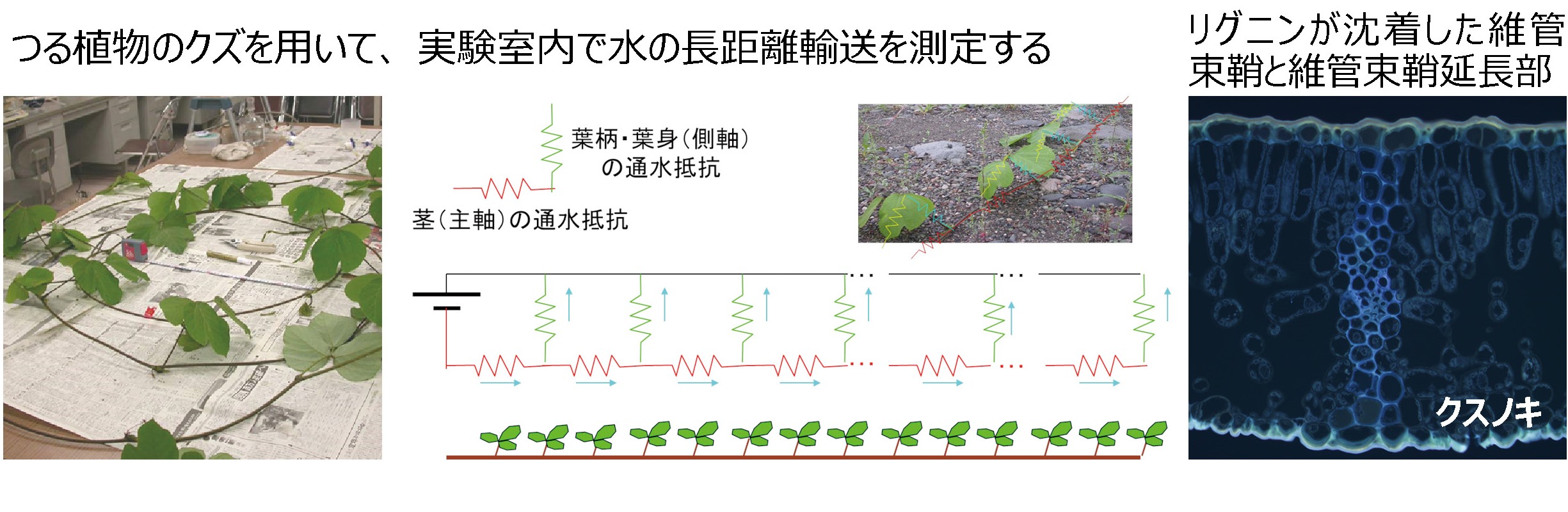
植物体内の均等な流れを調べる クズを用いた長距離輸送の研究の様子と輸送途中での水漏れを防ぐ葉脈の形態
文献: Taneda H & Tateno M (2007)Funct Ecol 21:226-234; Taneda H & Tateno M (2011)Tree Physiol 31:782-794; Taneda et al.(2016) Tree Physiol 36:1272-1282; Ohtsuka et al. (2017)Plant, Cell & Envirn 41:342-353
亜高山帯に分布する針葉樹では、冬の間、土壌や茎の凍結のために針葉への水の供給が止まります。健全な水分状態を保って冬を越すためには、針葉からの水の損失を小さく抑える必要があります。私たちは、針葉のクチクラや外表ワックスの結晶などの表面構造と水損失速度との関係を定量化しています。こうした解析から、越冬のために必要なクチクラやワックスの量や組成の解明を目指しています。
また、花弁を使って、花弁表面の撥水性と表皮細胞やクチクラのかたちの関係についても研究を行っています。花弁の表面構造はとても多様です。こうした多様な微細構造による撥水性への寄与や、この微細構造が生育環境によって変わるのかなど物理学的、生態学的な視点から解析を行っています。

針葉(シラビソ)のクチクラと花弁のクチクラ
文献: Taneda H et al. (2015) Ann Bot 115:923-937; Watanabe-Taneda A & Taneda H (2019) Flora 257:151417
*こちらのサイトで本郷キャンパスで採取した花弁の形態をまとめました。ご参照ください。
● 葉のサイズに合わせた葉脈の輸送システムの発生機構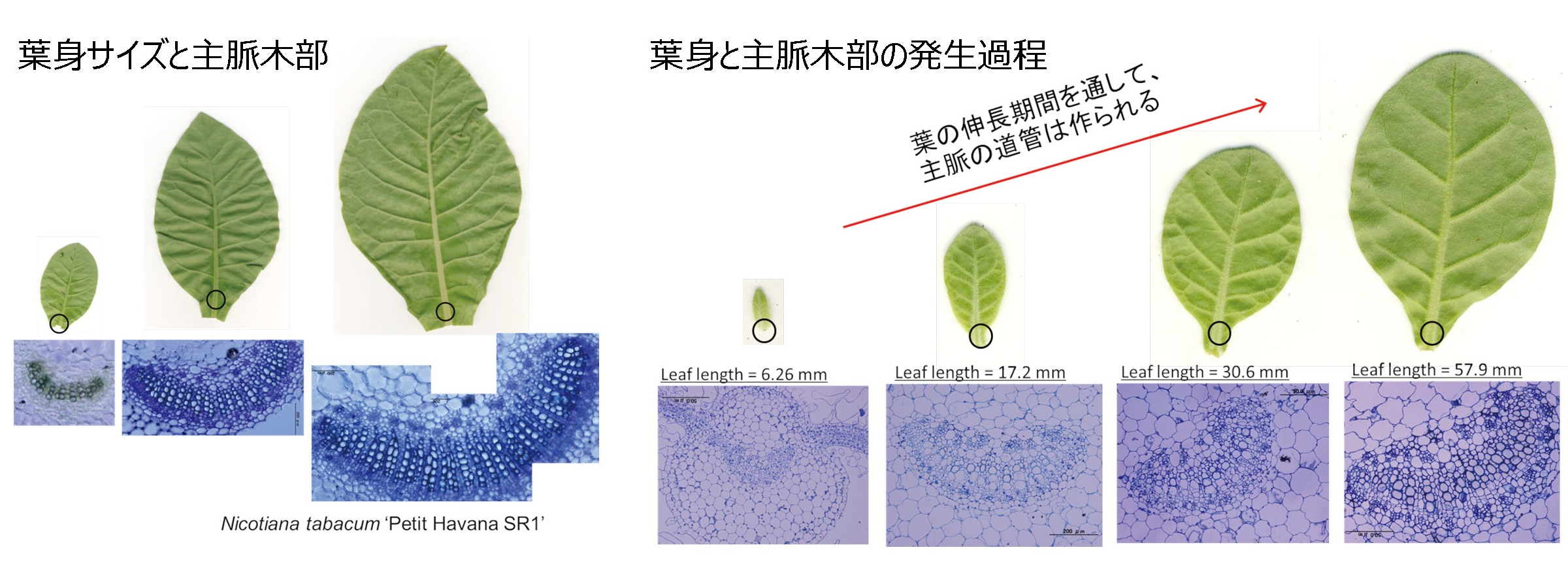
タバコにおける葉のサイズと木部の形態の関係
文献: Taneda H & Terashima I (2012) Ann Bot 110:35-45
私たちは木部閉塞に対する植物の生存戦略に興味を持ち、砂漠などの乾燥環境や冷温帯や亜高山帯といった寒冷環境への適応と木部閉塞への植物の応答についての研究を行ってきました。その結果、木部閉塞を起こしやすい茎の部分を冬に枯らすことで木部閉塞を回避したり、木部閉塞が起きにくい高い抵抗性を持つ種だけが寒冷地に分布していたり、木部閉塞の起きた道管や仮道管に水を再充填して通水能力を回復させたりといった、多様な生存戦略を明らかにしています。
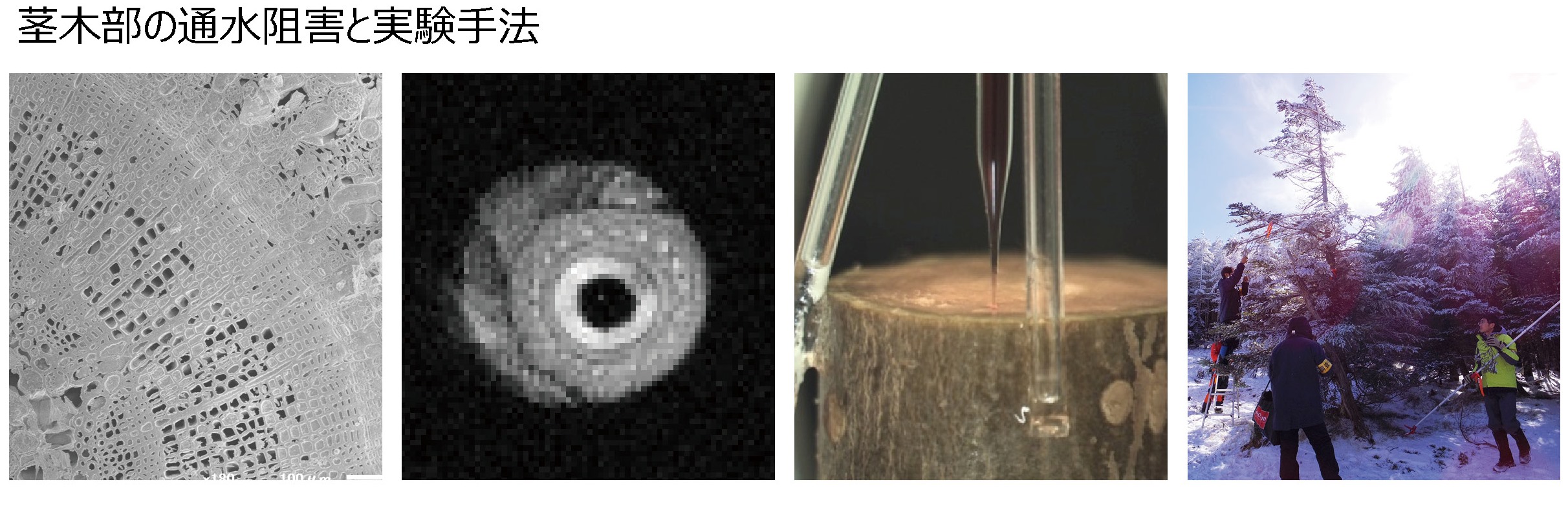
木部閉塞現象とその解析方法 左から、cryo-SEM法による木部水分布の可視化、MRI法による非破壊的な木部水分布の可視化、単一道管レベルでの還流法、枝の採取風景
文献: Taneda H & Tateno M (2005) Tree Physiol 25:299-306; Li Y et al. (2008) New Phytol 117:558-568; Taneda H & Sperry JS (2008) Tree Physiol 28:1641-1651; Sperry et al. (2012) Plant Cell & Envirn 35:601-610
これまでの私たちの研究から、冬の強風によって針葉や枝が枯死することが、生産性を低下させて個体の衰退するきっかけを作っていることがわかってきました。現在は、樹液流速の測定や年輪解析を通して、どのような速度で個体が衰退するのかを定量的に明らかにする予定です。また、衰退している個体がどのようなストレスを受けているのかを発現している遺伝子を調べて解析しています。

北八ヶ岳の縞枯れ現象と解析方法
文献: 種子田 (2018) 遺伝 72:57-62; Ogasa et al.(2019) Tree Physiol 39:1725-1735
北海道に生息するトドマツ(マツ科モミ属)は、広い標高域(200 m-1600 m)に分布しており、生息する標高環境に局所適応したエコタイプが確認されています。私たちは、東京大学農学部付属北海道演習林の天然林や共通圃場での植栽を用いて、局所適応している形態やゲノムワイドな相関解析 (GWAS) から複数の形質で産地標高と関連する遺伝子座の検出を試みています。こうした解析を突き詰めることで、局所適応のメカニズムの解明を目指しています。

北海道演習林と測定した形質の例
文献: Taneda et al. (2020) Trees 34:507-520