植物生態学研究室のメンバーのページへ
蜂谷卓士の研究内容 |
(研究A):高温下における呼吸速度増加メカニズムの解析Hachiya et al. (2007) Plant, Cell and Environment, vol. 30, p. 1269-1283 高温で栽培された植物では、しばしば組織の呼吸速度が増加する。この原因の一つは、タンパク質の代謝回転にかかるエネルギーコストの増加によると推測されてきた(Amthor 2000)。しかし、実証例はなかった。高温下における呼吸速度の増加(炭素のloss)は、植物の生産性を低下させる。したがって、そのメカニズムを知ることは重要な課題である。研究Aでは、タンパク質の代謝回転にかかるエネルギーコストの、高温下の呼吸速度増加への寄与を実測した。栽培温度の変化は、植物器官の発生速度に影響するため、植物器官の一生にわたるエネルギーコストを定量することにより、測定値の信頼性を高めた。実験材料として、単純なエネルギー収支を示す非光合成器官であり、寿命が短く一生にわたる解析に適したペチュニア花弁を用いた。タンパク質の代謝回転にかかるエネルギーコストは、タンパク質合成阻害剤添加時の呼吸速度の低下量から計算した(Noguchi et al. 2001)。その結果、高温下における呼吸速度の増加分のうち、45%がタンパク質の代謝回転にかかるエネルギーコストの増加によるものだった。また、高温下では、花弁のバイオマスが減少した。これは、タンパク質の代謝回転にかかるエネルギーコストの増加によって、成長に用いられる炭素が減少したことが一因と考えられる。研究Aの結果は、地球温暖化に伴う呼吸速度の増加に、タンパク質の代謝回転にかかるエネルギーコストの増加が大きく寄与することを示唆する。 |
(研究B):炭水化物の蓄積が成長呼吸係数(成長効率の指標)に及ぼす影響の解析Hachiya and Noguchi (2008) Physiologia Plantarum, vol. 134, p. 293-302 単位乾燥重量のバイオマスの合成に必要なエネルギーコストは成長呼吸係数と呼ばれ、植物種間の成長効率の差を議論する際の指標とされる。エネルギー代謝の分野において、「成長」とは炭水化物(基質)からバイオマスが合成されるプロセスと定義される(Amthor 2000)。したがって、炭水化物の蓄積は「成長」ではなく、エネルギーコストはほとんどかからない。一方、炭水化物含有率は乾燥重量の数%から数十%を占めるため、炭水化物の蓄積に伴って、成長呼吸係数は計算上減少する。炭水化物含量は、ストレスや日周によって大きく変動するため、成長呼吸係数を精度高く算出するには、乾燥重量全体から炭水化物含量を除く必要がある。研究Bでは、ペチュニア花弁の成長呼吸係数の実測値や他種の文献値に基づいて、成長呼吸係数の種間差や生育環境による差をより緻密に議論するためには、成長呼吸係数から炭水化物の寄与を除くことが必須であることを実証した。 |
(研究C):アンモニア毒性の「症状」と「原因」を判別する実験系の構築Hachiya et al. (2012) Plant and Cell Physiology, vol. 53, p. 577-591, 将来の高CO2環境や、世界的に大きな面積を占める酸性土壌においては、植物による硝酸態窒素の利用が抑制されるため、アンモニア態窒素の重要度が増す(Bloom et al. 2010 Science)。しかし、過剰のアンモニア態窒素は、植物にアンモニア毒性(硝酸と比較して「地上部の成長量・クロロフィル含量と主根の伸長が低下する現象」)を引き起こすため、今後、植物のアンモニア毒性をいかに抑制しつつ、アンモニア態窒素を吸収・同化させるか、(つまり、アンモニア毒性に関する研究)が重要な課題となる。1970年代から現在に至るまで、アンモニア毒性特有の「症状」が数多く記述されてきたが、アンモニア毒性の「原因」は特定されていない(Britto and Kronzucker 2002)。したがって、アンモニア毒性の数ある「症状」から「原因」を判別できる実験系の構築が急務であった。
|
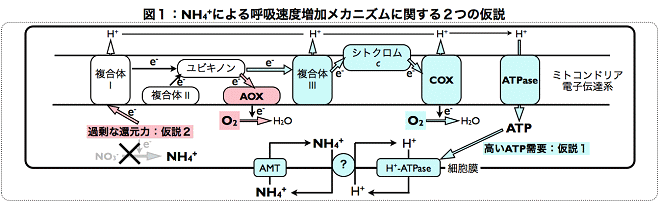
(研究D):アンモニア毒性における呼吸速度増加メカニズムの解析Hachiya et al. (2010) Plant, Cell and Environment, vol. 33, p. 1888-1897 多くの植物種において、アンモニアを窒素源として栽培したときの方が、硝酸を窒素源としたときよりも呼吸速度(酸素吸収速度)が高い(Britto and Kronzucker 2002)。この現象は、アンモニア毒性下でしばしば観察されるため、注目されてきた。アンモニアによる呼吸速度増加メカニズムとして、 (1) 細胞内からのアンモニアの排出に多量のATPが消費され、電子伝達系によるATP合成が高まること(図1 青色部:Britto et al. 2001)と、(2) 硝酸の還元反応に利用されなかった分の還元力が余り、電子伝達系で消費されること(図1 赤色部:Escobar et al. 2006)の2つが有力な仮説であった。仮説(2)では、ユビキノンから電子を直接酸素に受け渡すことのできるalternative oxidase (AOX)が重要な役割を担うと考えられている。AOXへの電子伝達はプロトンの輸送と共役しないため、ATP合成に寄与しないが、効率よく還元力を散逸できる。研究Dでは、8種類の組成の窒素源で栽培したシロイヌナズナを用いて、2つの仮説を比較・検証した。その結果、アンモニア依存的な呼吸速度の増加は組織内のアンモニアの蓄積によって説明され、硝酸の欠乏(硝酸の還元反応の低下)とは関係がなかった。また、aox変異株においても、野生株と同様のアンモニア依存的な呼吸速度の増加が観察され、他のさまざまな表現型にも違いは見られなかった。さらに、アンモニアを窒素源としたときには、ATPの合成と共役する経路(図1青色部)の発現や活性が誘導された。研究Dでは、2つの仮説を精巧な実験系によって初めて比較・検証し、仮説(1)を支持する明確な結論を得た。
|
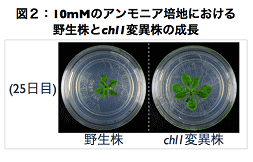
(研究E):硝酸センサーCHL1(Nitrate transporter 1.1)の硝酸非依存的な機能の解析Hachiya et al. (2011) Journal of Plant Research, vol. 124, p. 425-430 CHL1は硝酸の二重親和性輸送体としてだけでなく、硝酸のセンサーとして、様々な硝酸応答性の遺伝子発現を制御する(Ho et al. 2009 Cell)。ほとんどの先行研究では、CHL1の硝酸依存的な機能が解析されてきたが、一方で、硝酸非依存的な機能も示唆されていた。研究Eでは、CHL1のノックアウト変異株が高濃度のアンモニア(硝酸の無添加条件)に耐性を示すことを発見し(図2)、CHL1が硝酸非依存的な機能をもつことを実証した。このことは、CHL1を介したアンモニア感受性経路が存在することも意味する。CHL1を介した既知のシグナル伝達経路(硝酸に依存した経路)の変異株を用いた解析によって、アンモニア感受性経路は新奇の経路であることが明らかになった。
|
 論文1. Takushi Hachiya1, Ichiro Terashima1, Ko Noguchi1, Increase in respiratory cost at high growth temperature is attributed to high protein turnover cost in Petunia x hybrida petals, Plant, Cell and Environment, Blackwell Publishing Ltd., vol. 30, pp. 1269-1283, 2007. 2. Takushi Hachiya1, Ko Noguchi1, Effect of growth temperature and total non-structural carbohydrate accumulation on growth coefficient in Petunia x hybrida petals, Physiologia Plantarum, Blackwell Publishing Ltd., vol. 134, pp. 293-302, 2008. 3. Chihiro K. Watanabe1, Takushi Hachiya1, Ichiro Terashima1, Ko Noguchi1, The lack of alternative oxidase at low temperature leads to a disruption of the balance in carbon and nitrogen metabolism, and to an up-regulation of antioxidant defense systems in Arabidopsis thaliana leaves, Plant, Cell and Environment, Blackwell Publishing Ltd., vol. 31, pp. 1190-1202, 2008. 4. Takushi Hachiya1, Chihiro K. Watanabe1, Carolina Boom2, Danny Tholen2, Kentaro Takahara3, Maki Kawai-Yamada4, Hirofumi Uchimiya3, Yukifumi Uesono1, Ichiro Terashima1, Ko Noguchi1, Ammonium-dependent respiratory increase is dependent on the cytochrome pathway in Arabidopsis thaliana shoots, Plant, Cell and Environment, Blackwell Publishing Ltd., vol. 33, pp. 1888-1897, 2010. 5. Chihiro K. Watanabe1, Takushi Hachiya1, Kentaro Takahara3, Maki Kawai-Yamada4, Hirofumi Uchimiya3, Yukifumi Uesono1, Ichiro Terashima1, Ko Noguchi1, Effects of AOX1a deficiency on plant growth, gene expression of respiratory components and metabolic profile under low-nitrogen stress in Arabidopsis thaliana, Plant and Cell Physiology, Oxford University Press, vol. 51, pp. 810-822, 2010. 6. Keisuke Yoshida5, Chihiro K. Watanabe1, Takushi Hachiya1, Danny Tholen2, Masaru Shibata6, Ichiro Terashima1, Ko Noguchi1, Distinct responses of the mitochondrial respiratory chain to long- and short-term high-light environments in Arabidopsis thaliana, Plant, Cell and Environment, Blackwell Publishing Ltd., vol. 34, pp. 618-628, 2011. 7. Takushi Hachiya1, Yusuke Mizokami1, Kazunori Miyata1, Danny Tholen2, Chihiro K. Watanabe1, Ko Noguchi1, Evidence for a nitrate-independent function of the nitrate sensor NRT1.1 in Arabidopsis thaliana, Journal of Plant Research, Springer, vol. 124, pp. 425-430, 2011. 8. Takushi Hachiya1, Chihiro K. Watanabe1, Masaru Fujimoto1, Toshiki Ishikawa4, Kentaro Takahara3, Maki Kawai-Yamada4, Hirofumi Uchimiya3, Yukifumi Uesono1, Ichiro Terashima1, Ko Noguchi1, Nitrate addition alleviates ammonium toxicity without lessening ammonium accumulation, organic acid depletion and inorganic cation depletion in Arabidopsis thaliana shoots, Plant and Cell Physiology, Oxford University Press, vol. 53, pp. 577-591, 2012. [Editor-in-Chief’s choice article] |
総説等9. Takushi Hachiya1, Ko Noguchi1, Integrative response of plant mitochondrial electron transport chain to nitrogen source, Plant Cell Reports, Springer, vol. 30, pp. 195-204, 2011. 10. Takushi Hachiya1, Ko Noguchi1, Mutation of NRT1.1 enhances ammonium/low pH-tolerance in Arabidopsis thaliana, Plant, Signaling and Behavior, Landes Bioscience, vol. 6, pp. 706-708, 2011. |
脚注 1Department of Biological Sciences, Graduate School of Science, The University of Tokyo, Tokyo, Japan 2Plant Systems Biology Group, Partner Institute of Computational Biology, Shanghai, China 3Institute of Molecular and Cellular Biosciences, The University of Tokyo, Tokyo, Japan 4Department of Environmental Science and Human Engineering, Saitama University, Saitama, Japan 5Chemical Resources Laboratory, Tokyo Institute of Technology, Yokohama, Japan 6Faculty of Education, Yamaguchi University, Yamaguchi, Japan このページのトップへ 植物生態学研究室のページへ |