|
 日本語 / 日本語 /  English English
|

研究概要
(「これまでの研究成果」に関しては、見易さのため個々の文献を引用しておりません。全て業績リストに収録されていますので、そちらをご参照下さい。) ミツバチは高度な社会を形成して生活する真社会性昆虫であり、そのコロニーでは、個体間でさまざまな役割が分担されている。また、働き蜂は「尻振りダンス」を用いて巣仲間に餌場の位置(距離と情報)に関する情報を伝達する。こうした高度な行動や社会性の成立を可能とする分子的基盤を幾つかの側面から研究している。 (1) ミツバチの社会性行動と「尻振りダンス」の分子・神経的基盤の解析 【研究の背景】 ヒトの脳では異なる脳機能が大脳皮質の別々の領野により分担されている(「脳機能局在論」)。例えば言語能力に関わる領野としては、言語理解に関わるウェルニッケ野と音声発声に関わるブローカ野が存在する。もし、言語野選択的に発現する遺伝子が見つかれば、言語野の働きを理解する上で非常に有効な手掛かりになるだろう。しかし、そうした遺伝子は未だ見つかっていない。またサルは言語能力をもたないため、言語能力の研究のためのモデル生物は存在しない。 【研究の目的】 私たちは、ミツバチの社会性行動や「尻振りダンス(ダンス言語)」を産み出す脳機能を理解する上で、これと同じ研究戦略を採用している。つまり、(1)ミツバチの社会性行動や「尻振りダンス」に関わる脳領野を調べると共に、その領野選択的に発現する遺伝子を同定し、その働きを人為的に修飾することで、高次行動を産み出す脳の仕組みを調べようとしている。これまでの研究で次のことが明らかとなってきた。なお、特に断らない限り、ここでは「ミツバチ」は「セイヨウミツバチ(Apis mellifera L.)」を指す。 【これまでの研究成果】
ハチの仲間には、単独性から亜社会性(一時期、自分自身の子と一緒に生活する)、共同巣性(アパートのように、同世代のハチが同じ巣を使い、別々に子育てをする)、疑似社会性(「共同巣性」+共同で子育てをする)、半社会性(「疑似社会性」+カーストが存在する)、真社会性(「半社会性」+親世代と子世代が重複する)といった異なる社会性進化の段階にある種類が知られているが、こうした社会性の進化を可能にした分子・神経的機構は不明である。 私たちはこうしたハチ目昆虫の社会性進化は「新しい脳領野(遺伝子発現プロフィル)の獲得と、それによる新しい脳機能の獲得に基づく」のではないかと考えている。特にmKastを発現する「中間型」ケニヨン細胞は、採餌飛行をする亜社会性の段階で出現し、その後、尻振りダンスの基礎となった可能性が考えられる。今後はゲノム編集技術などを用いて、上記で提案した遺伝子機能や脳領野の機能を検証すると共に、異なる社会性進化の段階にあるハチ目昆虫での、脳領野や遺伝子発現プロフィルの比較を行う予定である。 (2) ミツバチの働き蜂の分業に伴う生理状態変化の分子機構の解析 【研究の背景と目的】 ミツバチでは雌が女王蜂と働き蜂にカースト分化し、さらに働き蜂は羽化後の日齢に伴って羽化後2?3週間は育児(ローヤルゼリーを口から分泌し、幼虫に与える)や巣の掃除に従事し、門番を経て、羽化後2?3週間以降は花蜜や花粉の採集(採餌)へと分業(「例差分業」)する。この時、頭部分泌腺である下咽頭腺は育児蜂では発達し、ローヤルゼリー(RJ)を分泌するが採餌蜂では退縮し、花蜜のスクロースをグルコースとフルクトースに分解するα-グルコシダーゼの活性が検出されるようになることが知られていた。私たちは、働き蜂の分業において行動と生理状態が協調的に変化する仕組みに興味をもち、次のようなことを明らかにしてきた。 【これまでの研究成果】
個々の働き蜂がもつ生理状態(下咽頭腺の遺伝子発現プロフィル)は、働き蜂の日齢ではなく、その役割に応じて協調的に調節されると考えられる。これまで、働き蜂の分業に働くホルモンとしてはJHのみが知られてきたが、少なくとも下咽頭腺の機能胆管には、JHとエクダイソンが協調的に働くことが示唆された。(1)の⑥⑦で述べたように、働き蜂の脳でエクダイソンが合成され、小型ケニヨン細胞にEcRが選択的に発現することを考え合わせると、エクダイソンが脳(小型ケニヨン細胞)と下咽頭腺の細胞機能を協調的に制御する可能性が考えられる。 齢差分業はハチ目昆虫でもミツバチなど、ごく限られた種類にのみ見られる行動形質である。元来、変態のためのホルモン制御を、ミツバチは働き蜂の分業のために「再利用」したのかも知れない。今後は(1)の課題と同様に、ゲノム編集法を用いて、それぞれの遺伝子機能を実証する予定である。特に下咽頭腺や他の分泌腺の生理状態を人為的に変調させた時、その働き蜂がどう振る舞うかは分業を成立させた分子機構を考える上で興味深い課題ではないかと考えている。 (図1) 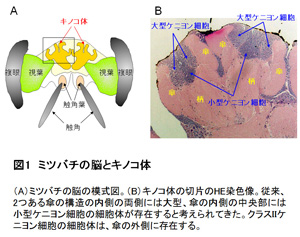 (図2) 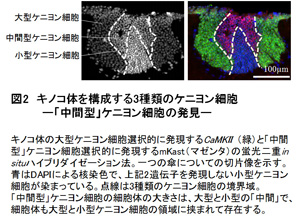 (図3) 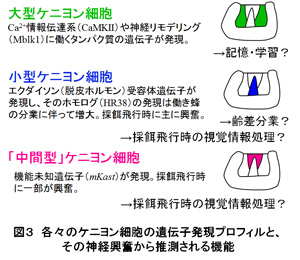 【理学系研究科からのプレスリリース】 1. 採餌飛行をするミツバチの脳で活動する新規な神経細胞「中間型ケニヨン細胞」の発見 (金子九美・久保健雄) http://www.s.u-tokyo.ac.jp/ja/press/2013/41.html http://www.u-tokyo.ac.jp/ja/utokyo-research/research-news/discovery-of-novel-honeybee-brain-neurons-active-during-foraging-flight/ 2. ニホンミツバチの攻撃行動(熱殺蜂球形成)における脳の活動を解明 (宇賀神篤・木矢剛智・國枝武和・小野正人・吉田忠晴・久保健雄) http://www.s.u-tokyo.ac.jp/ja/press/2012/11.html 3. 脳の配線ミスを生後の発達期に修正する新たなメカニズムの発見 (林悠・飯野雄一・久保健雄) http://www.s.u-tokyo.ac.jp/ja/press/2009/14.html
失われた器官や付属肢を再形成する“再生能”は多くの動物種が有しているが、その程度は動物の種や器官、発生ステージなどにより様々に異なる。例えば、プラナリアは一個体を数十の断片に切断しても、それぞれの断片から個体が再生するという非常に高い再生能を持つ。無尾両生類(カエルなど)では、幼生(オタマジャクシ)の尾を切断すると、表皮・血管・筋肉はもとより脊索・脊髄も再形成されるが、成体であるカエルの四肢は切断された場合、指を欠いた“スパイク”と呼ばれる構造が再形成されるに留まる。一方、哺乳類においては肝臓を除き高度な再生能を示す器官は存在しない。このように、動物の再生能が多様である理由、つまり何が再生能を規定する要因かについては、今なお明らかではない。 当研究室では、アフリカツメガエル(Xenopus laevis)幼生尾再生を実験系に用い、生体防御と再生の関連について研究を進めている。アフリカツメガエル幼生の尾は前述のように高い再生能をもっているが、発生過程の特定の一時期(受精後約1~3週間)においては尾再生がおきない(再生不応期)ことが知られていた(図1)。当研究室ではこれまで、再生不応期の個体を薬理学的(NF-κB阻害剤・免疫抑制剤)に免疫抑制した場合、また各種免疫細胞への分化に必要な転写因子(PU.1)のノックダウンにより免疫細胞を減少させた場合に有意に再生能が回復することを見出し(図2)、免疫応答はツメガエル幼生尾の再生能を規定する大きな要因の一つであることを提唱した(図3)。現在、この免疫応答の実体解明(どの種の免疫細胞が、どのように再生を阻害するのか)に向けて研究を行っている。 現時点の作業仮説としては、不応期型の免疫応答は自己抗原を認識する自己反応性の免疫細胞によるものとし、その自己抗原は再生芽(尾切断後に形成される増殖細胞集団)を形成する細胞において発現していると考え、これら攻撃実体の同定とそれが認識する抗原の探索を行っている。またこの探索過程で、再生芽増殖細胞において選択的に発現している遺伝子群が同定されており、これら遺伝子群が再生芽増殖細胞の維持・再生尾形成に関与している可能性についても検討を行っている。 (プレスリリース:インターロイキン11は、オタマジャクシの尾再生芽の未分化細胞の誘導・維持に働く—器官再生の最初期段階の人為的再現に成功—) 私たちは、この“免疫応答は再生能を規定する一因である”という概念は哺乳類へ敷衍できるのではないかと考えている。哺乳類では、深い傷を負うと炎症が生じ治癒した後には傷跡が残るが、免疫系の未発達な胎児期では重い炎症は起こらず、傷跡が残らないで治癒する。前述のPU.1を欠損するマウスは様々な免疫細胞を欠くが、このマウスでは成体になっても傷跡が残らない治癒が起こる。PU.1依存に発生する免疫細胞は、両生類と哺乳類に共通して、器官再生や創傷治癒の能力を低下させる要因となっている可能性がある。ほかにも、胸腺を欠くヌードマウスや、T細胞による細胞傷害に関わるFasに変異を持つMRLlprマウスといった免疫系に異常を持つマウス系統はしばしば成体でも傷跡を残さない治癒が起こる。このように、哺乳類においても、免疫応答は組織の再構築を阻害する要因となり得る。哺乳類において免疫抑制が直ちに再生能の賦活化を引き起こすとは考えにくいが、器官再生を実現するうえでの必要条件の一つである可能性は高い。私たちのツメガエル幼生尾再生不応期の研究においてその攻撃実体と抗原が同定されれば、組織・器官損傷後にその抗原を免疫系から保護することで、再生能の人為的賦活化が可能になるかもしれない。このような、哺乳類での器官再生実現へ向けた研究展開ができればと考えている。 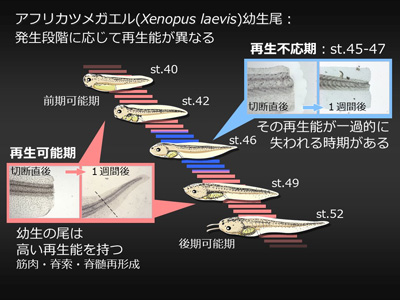 (図1)アフリカツメガエル幼生尾は発生時期に応じて再生能が異なる。元々再生が可能であった(前期可能期:受精後4~5日)ものが、その再生能を一過的に失い(不応期:6日~約3週間)、その後再び再生するようになる(後期可能期:約3週間以降)。 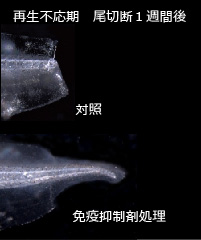 (図2)再生不応期に免疫抑制すると再生能が回復する。図は免疫抑制剤(FK506)処理した不応期個体尾切断1週間後。対照個体は再生していない。 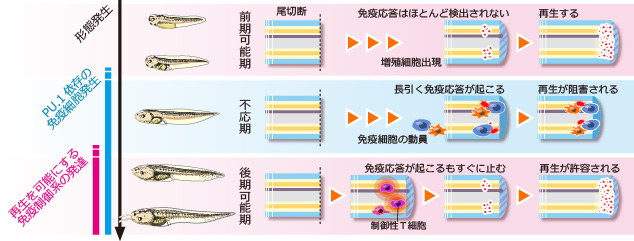 (図3)尾の再生能の変遷と免疫発生・応答のモデル図。前期可能期では免疫系がまだ発生していないため顕著な免疫応答は起きず、尾は再生する。不応期ではPU.1依存の免疫細胞が出現し始め、これらが尾の再生に阻害的に作用するような“不応期型の免疫応答”を起こす。その後後期可能期までに免疫制御系が発達し尾切断時に不応期型の免疫応答が抑えられることで再生に寛容になる。
クマムシ研究グループの詳細ページはこちら 地球上には実に様々な動物たちが生息しているが、中には我々人間が及びもつかないような能力を持つものも少なくない。こうした中で、最も極限 的な環境ストレスに耐えると言われている動物がクマムシ類である。陸生クマムシの多くは乾燥耐性を持ち、周囲が乾燥すると脱水して縮まり乾眠 と言われる状態に移行する。この状態では水をほぼ完全に失っており、生命活動は一切見られない。しかし、死んだわけではなく水を与えると速や かに生命活動を再開する。乾眠状態では、驚異的な極限環境(-273~151度、真空~75,000気圧、ヒトの致死量の1000倍の放射線)に耐性を示すこ とが知られている。 クマムシの耐性能力は約230年前から知られているにも関わらず、その分子機構は全くといって良いほど分かっていない。私たちはクマムシの耐 性能力のメカニズムの解明を目的として、乾燥耐性に関わる実働因子の同定と解析を進めている。実働分子であるタンパク質に着目したプロテオミ クス解析(発現プロテオミクス、機能プロテオミクス)のほか、他研究室と共同でクマムシの全ゲノム解読およびトランスクリプトーム解析を進め ている。 クマムシの乾眠のメカニズムから、動物の細胞や組織を乾燥保存できる仕組みが明らかになれば、ヒトの医療や産業など応用面にも大きな影響を 及ぼすことが期待される。また、乾眠状態のクマムシが吸水して復活する過程は、生命活動を示さない物体から生命活動が始まる過程ともいえるこ とから、この過程で何が起きているかを調べることで、生命とは何かという根本的な疑問に迫れるのではないかと期待している。さらに研究が進め ば、人工的に生命活動を生み出すこともできるようになるかもしれない。
|